

Summary
ここでは、機能性分子複合体を追跡し、定量、検出できるようにするために細菌細胞を生きて単一分子蛍光顕微鏡を実行するためのプロトコルを示す。
Abstract
生きた細胞のメカニズムへの完全な洞察は、細胞レベルでの事象を惹起し、ダイレクトキープロセスを調べることによってのみ達成することができます。生物系のせん断複雑さをこれまでには、代わりに相対的に原油バルクアンサンブル平均測定を使用して単一システムの研究に焦点を当て、あまりにも要求される正確な単一分子実験を引き起こしている。しかし、多くの重要なプロセスは一つのレベルで生きている細胞や少数の分子で起こる、アンサンブルの測定値は、一般的にこれらのイベントの確率と異種の性質を覆い隠す。ここでは、高度な光学顕微鏡と我々は、単一の分子とどのように我々は生物学的機械の機能に分子複合体内動態を観察できるの精度への単一の生きた細菌細胞内のタンパク質を監視する方法を示す分析画像解析ツールを使用して。テクニックは、生理学的に直接関連しています。彼らが検討されて生体試料に最小限の摂動と非侵襲的であり、完全に生体を構成する物質の調査、生物物理学の他の単一分子のアプローチが容易に利用できない機能に同調されています。さらに、生物学的標本は、すべて自然に発生するよりもはるかに多くのタンパク質を生成するための、より一般的ですが、理想的なアプローチとは対照的に変更されていない細胞株("ゲノムエンコード")とほとんど同一のレベルで、少なくとも蛍光タグタンパク質を生成する研究("発現プラスミド")。このように、調査される実際の生物学的サンプルは、自然の生物にかなり近いですので、実際の生理学的プロセスへの観察は、より関連性の高い。
Protocol
- この手順を開始するには、 大腸菌の細菌細胞を発現する蛍光蛋白質の凍結ストックの50μlを最初に解凍し、午前中は37℃で一晩5 mlのLB培養培地で振盪しながら好気的に栽培されている、この飽和文化の50μlを抽出し、 、最小のM63グルコースの培地へと継代培養4〜6時間、30℃でインキュベートする。ここでは、電子輸送を表現するそのうちの一つ、2つの異なる細胞株を、使用例を示すGFP、GFPと融合した細菌べん毛モーターに関与するタンパク質を発現する他に融合されたチトクローム。
- 細胞は固定化のサンプルとしてそれらを表示する場合のいずれかに成長のサブカルチャーから直接採取したり、それらは"テザー"の条件で表示する場合細菌の鞭毛を切り詰めるために剪断することができます。
- せん断は、無菌配管で結合された2つの滅菌注射器で構成されるデバイスに典型的にはサブカルチャーの1〜5ミリリットルを置くことが含まれます。せん断は、狭いチューブを通して文化をプッシュするごとにシリンジポンプを押し込んで交互に行われます。これは、必要なせん断の程度に応じ、50〜100倍が行われる。文化は、鞭毛のフラグメントを削除するには、最少培地で再懸濁している細胞をペレットに遠心分離されています。
- 私たちは、その後、その後、脱イオン水とエタノールで徹底的に洗浄し、少なくとも1時間空気中で乾燥するまま、20分間エタノールにKOHの飽和溶液に浸漬することにより、BK7ガラス製カバースリップの洗浄準備
- 我々は、顕微鏡で細胞を収容するための簡単なフローセルを構築する。これは、BK7ガラスの顕微鏡スライド上にパラフィングリースの描画線を含み、その上にきれいにcoverlipsのいずれかを配置し、ピンセットで軽く押し下げて、トンネルサンドイッチを作成、5〜10μlののフローセルのボリュームを与える。
- 固定化細胞を観察するために我々は、ポリ- L -リジンの0.1%w / v溶液を注射することにより、フローセルを入力し、それは少なくとも1分間室温でインキュベートすることができます。我々は、その後、フローセルの一方の端からメディアを注入し、他からティッシュペーパーでウィッキングで100μlの最少培地を用いて洗い流す。
- 私たちは、その後、カバーガラスの表面をマークする最少培地で200nmの直径ラテックス微小球(Polysciences)の1:500希釈の20μlを通過ウィク。フローセルは、カバースリップは、下向きにシンプルな湿度チャンバーで表面の明確なプラットフォーム上に配置し、5分間室温でインキュベートされるように反転されます。非結合ビーズを100μlの最小限のメディアを介して吸い上げによって洗い流されている。
- 我々がつながれた細胞を観察したい場合は我々は、ポリ- L -リジンのステップを省略し、代わりに5 mg / mlの抗フラジェリン抗体とフローセルを埋める。フローセルは、10分間加湿チャンバー内の場所ですし、発散性に優れたによってによってフラッシュされます。
- 細胞培養液20μlの固定化細胞を見に係留細胞または断片化していない試料を観察する場合のどちらか剪断されたサンプルを使用して、フローセル通過して邪悪です。
- フローセルが反転し、20分間加湿チャンバーに配置されます。非結合細胞を次いで100μlの最小限のメディアを通して吸い上げによって洗い流されている。
- 液浸オイルのドロップはカバースリップの上面の中央に配置され、フローセルは、高開口数対物レンズとの光学的接触をする、カスタムビルドの蛍光顕微鏡の試料ホルダーにに軽く置かれます。
- 顕微鏡電子乗算カメラがオンになり、カメラは-70℃に冷却するように設定され、ソフトウェアは、最初のゲインコントロールをdisenabling、フレーム転送モードで25Hzの典型的なフレームレートで画像を取得するために設定されていますカメラ。
- 明視野照明のスイッチがオンになっていると画像がそのカバーの表面に平行に、その長軸と立ち往生しているに基づいて撮像するセルの適切なセルまたはグループを選択し、フォーカスに取り込まれます。フォーカスが細かくカバーの表面に付着した200nmのラテックスビーズをジャストフォーカスになっていることを保証するために調整されます。
- イメージシーケンスは、細胞体の輪郭を記録するために明に取得されます。明視野照明はオフになり、カメラのゲインが最大に有効になっています。
- 内部全反射蛍光(または反射)、適切な波長以上のレーザービームを(ここで、緑色蛍光タンパク質の励起のために、我々は473 nmのレーザーを使用)を使用して標準的な取得のための後焦点面に集束されるようにあらかじめ設定されています対物レンズが、細胞のサンプルに蛍光励起のためのエバネッセント場を生成する光軸から横方向にずれる。
- カメラの取得が開始され、レーザーのシャッターには、細菌内で蛍光タンパク質を励起するために開かれます。買収のレーザーの強さと速度のパラメータは異なる値を試すことで調査中の特定の生物学的システム向けに最適化する必要がありますが、典型的な範囲R細胞膜におけるモバイルタンパク質複合体を研究するelevantは50から40 msのフレームあたりの露光時間で、直径〜30μm程度の円形の励起領域の上に1から10 mWのレーザパワーです。サンプルは通常、〜10秒間、photobleachedまで点灯されています
- このプロトコルは、両方のセルの本体は、カバースリップと鞭毛スタブとの間の、そして堅くカバーの表面に固定されている固定化細胞のための結合点を中心に回転されるテザーセルに使用することができます。
- 複合体の高コピー数を持ついくつかの細胞株は、前の全反射イメージングにセルの一方の極の初期回折限界の集光漂白剤の恩恵を受ける。この漂白剤は、(またはFRAP)光退色後蛍光回復で使用されているものと同等です。私たちのカスタム顕微鏡を使用して、励起レーザ光の一部は、FRAP型漂白に使用する第2の独立したパスにオフ供給することができます。通常は1 - 10mWのレーザパワーは、範囲10から300ミリ秒の典型的な漂白時間は、使用されています。その後、より簡単に可視化するためにその領域に拡散させる個々の複合体を可能にする細胞の漂白ゾーンではるかに高いイメージングコントラストでこの結果。
- "slimfield"と呼ばれる別の照明モードが採用されている細胞膜に比べてはるかに高速拡散細胞の細胞質で複合体を視覚化するための。ここで焦点レーザーは、横方向にちょうど単一のセルを含むように拡張されている。これは通常、ミリ秒のかなり高速のエクスポージャーを取ることができ、非常に強烈なフィールドを生成します。
- データ取得に続いて、画像はカスタム作成されたソフトウェア(LabVIEW 8.5でコード化された)に供給される。これは自動的に通常、数ナノメートルの精度に細胞内の蛍光スポットの位置を検出し、その大きさと明るさを抽出します。追跡分子複合体の時間に対する光退色トレースはその後化学量論を推定するために使用されているbrightnessofは、単一の分子複合体を構成する方法多くの個々の蛍光タンパク質、すなわち。
代表的な結果:
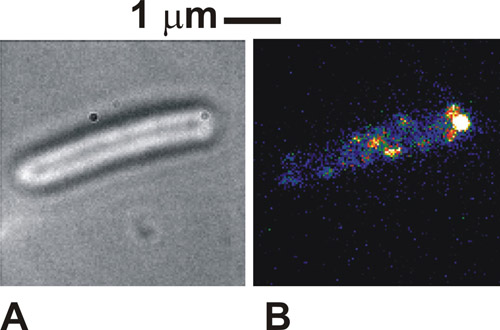
プロトコルが正しく行われている場合、明視野で見る細胞の画像は白/グレーの細胞体(図1a)に対して暗細胞体の周囲で、非常に区別される。蛍光を使用して固定化細胞では、我々は、幅で、通常は250から300 nmの(図1b)の別個のスポットの強度を、見ることができます。健康、テザー細胞は、明視野像におけるテザーの添付ファイルのポイントを中心に回転するように見られます。蛍光励起の下で我々の場合におけるいくつかの分子複合体はまた、べん毛モーターを持つタグタンパク質の局在を示す、添付ファイルの時点で見られるかもしれません。これらのスポットは、個々の分子の複合体であると見られるそれらの数は、使用される照明モードに依存し、実際に一度に細胞内でどのように多くの複合体の提示されています。スポットの移動度は、研究対象の特定の生物学的システムに依存します。スポットの密度が最初は非常に高い場合は、ここで使用ラベル付けシトクロムの場合と同様に、初期FRAPの漂白を行うと、画像のコントラストを向上させることができます。

図1。細菌の鞭毛モーターに関与することが知られている緑色蛍光タンパク質(GFP)に融合タンパク質を発現する固定化大腸菌の細胞のための(A)明視野と(B)全反射の画像(疑似カラー)。してくださいここをクリックして図1の拡大バージョンを参照すること。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
これは、べん毛モーターの機能を損なうおそれがあるので注意が、つながれた細菌を調べるための細胞"せん断を介して"しないように注意する必要があります。彼らは酸素枯渇になる可能性があるので、顕微鏡スライド上に、1時間に1回よりもはるかに長いためにセルを使用することが重要です。かなりの最適化は、最適な顕微鏡の撮像条件が検討され、特定の生物学的システムへの仕出し料理見つけるために必要となる場合があります。それはあなたの特定の顕微鏡システムに必要な正しい強度レーザーの励起を確認するために単独で精製GFPを用いたイメージングを試みることが賢明かもしれない。
Subscription Required. Please recommend JoVE to your librarian.
Acknowledgments
我々は教授ジュディスアーミテージ(オックスフォード大学、英国)と教授コンラッドMullineaux(ロンドンのクイーンメアリー大学、英国)のグループから、菌株の種類の寄付を認める。 IMDは、共同で生化学の部門(オックスフォード大学)とOCISBによって資金を供給され、ARは、工学物理科学研究評議会(EPSRC)DTCの学生の身分によって資金を供給され、NDは、バイオテクノロジーおよび生物科学研究会議(BBSRC)から資金提供され、MCLは王立協会大学研究フェローシップによって資金を供給。
References
- Leake, M. C., Chandler, J. H., Wadhams, G. H., Bai, F., Berry, R. M., Armitage, J. P. Stoichiometry and turnover in single, functioning membrane protein complexes. Nature. 443, 355-358 (2006).
- Leake, M. C., Greene, N. P., Godun, R. M., Granjon, T., Buchanan, G., Chen, S., Berry, R. M., Palmer, T., Berks, B. C. Variable stoichiometry of the TatA component of the twin-arginine protein transport system observed by in vivo single-molecule imaging. Proc Natl Acad Sci U S A. 105, 15376-15381 (2008).
- Leake, M. C., Wilson, D., Gautel, M., Simmons, R. M., M, R. The elasticity of single titin molecules using a two-bead optical tweezers assay. Biophys. J. 87, 1112-1135 (2004).
- Leake, M. C., Wilson, D., Bullard, B., Simmons, R. M. The elasticity of single kettin molecules using a two-bead laser-tweezers assay. FEBS Lett. , 535-555 (2003).
- Lenn, T., Leake, M. C., Mullineaux, C. W. In vivo clustering and dynamics of cytochrome bd complexes in the Escherichia coli plasma membrane. Mol. Microbiol. 70, 1397-1407 (2008).
- Lenn, T., Leake, M. C., Mullineaux, C. W. Are Escherichia coli OXPHOS complexes concentrated in specialised zones within the plasma membrane. Biochem. Soc. Trans. 36, 1032-1036 (2008).
- Lo, C. J., Leake, M. C., Pilizota, T., Berry, R. M. Single-cell measurements of Membrane Potential, Sodium-Motive Force and Flagellar Motor Speed in Escherichia coli. Biophys. 93, 294-302 (2007).
- Lo, C. J., Leake, M. C., Berry, R. M. Fluorescence measurement of intracellular sodium concentration in single Escherichia coli cells. Biophys. J. 90, 357-3565 (2006).

{kind=link}