


Summary
Dit protocol beschrijft een niet-invasieve methode om S-fase cellen efficiënt te identificeren voor downstream microscopiestudies, zoals het meten van DNA-reparatie eiwitrekrutering door lasermicrobestraling.
Abstract
DNA-schadeherstel handhaaft de genetische integriteit van cellen in een zeer reactieve omgeving. Cellen kunnen verschillende soorten DNA-schade accumuleren als gevolg van zowel endogene als exogene bronnen zoals metabole activiteiten of UV-straling. Zonder DNA-reparatie wordt de genetische code van de cel aangetast, waardoor de structuren en functies van eiwitten worden ondermijnd en mogelijk ziekte wordt veroorzaakt.
Het begrijpen van de spatiotemporale dynamiek van de verschillende DNA-reparatieroutes in verschillende celcyclusfasen is cruciaal op het gebied van DNA-schadeherstel. De huidige fluorescerende microscopietechnieken bieden geweldige hulpmiddelen om de rekruteringskitiek van verschillende reparatie-eiwitten na DNA-schade-inductie te meten. DNA-synthese tijdens de S-fase van de celcyclus is een eigenaardig punt in het lot van cellen met betrekking tot DNA-reparatie. Het biedt een uniek venster om het hele genoom te screenen op fouten. Tegelijkertijd vormen DNA-synthesefouten ook een bedreiging voor de DNA-integriteit die niet wordt aangetroffen in niet-delende cellen. Daarom verschillen DNA-reparatieprocessen aanzienlijk in de S-fase in vergelijking met andere fasen van de celcyclus, en die verschillen zijn slecht begrepen.
Het volgende protocol beschrijft de voorbereiding van cellijnen en de meting van de dynamiek van DNA-reparatie-eiwitten in S-fase op lokaal geïnduceerde DNA-schadeplaatsen, met behulp van een laserscannende confocale microscoop uitgerust met een 405 nm laserlijn. Tagged PCNA (met mPlum) wordt gebruikt als een celcyclusmarker in combinatie met een AcGFP-gelabeld reparatie-eiwit van belang (d.w.z. EXO1b) om de DNA-schaderekrutering in de S-fase te meten.
Introduction
Verschillende DNA-reparatieroutes zijn geëvolueerd om de verschillende soorten DNA-laesies aan te pakken die in cellen kunnen ontstaan, die allemaal sterk gereguleerd zijn in zowel ruimte als tijd. Een van de meest kwetsbare periodes van de celcyclus is de S-fase, wanneer DNA-synthese plaatsvindt. Hoewel proliferatie fundamenteel is voor het leven, biedt het ook een grote uitdaging. Cellen moeten zorgen voor een getrouwe replicatie van hun genoom om te voorkomen dat mutaties worden doorgegeven aan toekomstige generaties. Bijgevolg biedt proliferatie een therapeutisch interventiepunt dat is gebruikt voor de ontwikkeling van therapeutische benaderingen op het gebied van oncologie.
Alle belangrijke technieken die worden gebruikt voor het bestuderen van eiwitrekrutering bij DNA-laesies hebben hun sterke punten en beperkingen. Micro-bestraling heeft een betere ruimtelijke en temporele resolutie1 dan de meeste alternatieve methoden zoals immunofluorescente beeldvorming van ioniserende straling-geïnduceerde foci (IRIF), chromatine-immunoprecipitatie (ChIP) of biochemische fractionering. Microbestraling doorspekt echter de robuustheid van de bovengenoemde technieken die een groot aantal cellen tegelijkertijd kunnen bemonsteren.
Om DNA-reparatie in S-fase te onderzoeken, moet men S-fasecellen kunnen onderscheiden in een asynchrone celkweekpopulatie. Er zijn veel bekende methoden om dit aan te pakken, waarbij de synchronisatie van cellen of visualisatie van de verschillende celcyclusfasen betrokken zijn. Beide benaderingen brengen echter aanzienlijke uitdagingen en mogelijke artefacten met zich mee. Chemische synchronisatiemethoden die veel worden gebruikt om cellen in de vroege S-fase te verrijken (bijv. Dubbele thymidineblok-, aphidicolin- en hydroxyurea-behandeling) bereiken synchronisatie door de inductie van replicatiestress en uiteindelijk DNA-schade zelf. Dit beperkt het gebruik van deze methoden om DNA-reparatieprocessen in S fase 2 tebestuderen. Synchronisatie door serumuithongering en -afgifte is slechts van toepassing op een beperkt aantal cellijnen, grotendeels met uitzondering van kankercellijnen die minder afhankelijk zijn van groeifactoren voor celcyclusprogressie in vergelijking met niet-getransformeerde cellijnen. Het Fluorescence Ubiquitin Cell Cycle Indicator (FUCCI) -systeem is een bijzonder nuttig hulpmiddel om de celcyclus te bestuderen, maar het heeft een fundamentele beperking bij het onderscheiden tussen S- en G2-celcyclusfasen3.
Hier wordt aangetoond dat het gebruik van fluorescerend gelabeld PCNA als een niet-invasieve marker voor S-fase de nadelen van chemische celcyclussynchronisatiemethoden beperkt, terwijl meer specificiteit en flexibiliteit mogelijk is dan het FUCCI-systeem. Als een enkele marker kan PCNA niet alleen S-fasecellen in een asynchrone populatie markeren, maar het kan ook de exacte progressie van cellen binnen de S-fase (d.w.z. vroege, midden- of late S-fase) laten zien4. Lage expressieniveaus van exogene, gelabelde PCNA zorgen voor minimale interferentie met zowel celcyclusprogressie als DNA-reparatieprocessen. Belangrijk is dat PCNA ook dient als een interne controle voor de juiste DNA-schade-inductie, omdat het betrokken is bij het herstel van verschillende DNA-laesies en wordt gerekruteerd naar lokaal geïnduceerde DNA-schadeplaatsen1,4.
De hier gepresenteerde experimenten laten zien hoe de rekruteringsdynamiek van EXO1b in S-fase kan worden gemeten en hoe dit wordt beïnvloed door de gevestigde PARP-remmer olaparib. EXO1b-nuclease-activiteit is relevant voor een breed scala aan DNA-reparatieroutes, waaronder mismatch-reparatie (MMR), nucleotide-excisieherstel (NER) en dubbelstrengsbreuk (DSB) -reparatie. In de S-fase speelt EXO1b een belangrijke rol bij homologe recombinatie (HR) door de vorming van 3' ssDNA-overhangen tijdens DNA-resectie5. EXO1b is verder betrokken bij DNA-replicatie met rollen in checkpoint-activering om vastgelopen DNA-vorken opnieuw op te starten, evenals primerverwijdering en Okazaki-fragmentrijping aan de achterblijvende streng tijdens strengverplaatsing in replicatie5. EXO1b-rekrutering naar beschadigde DNA-plaatsen wordt gereguleerd door de directe interactie met poly (ADP-ribose) (PAR)6,7. Vanwege de vele celcyclusspecifieke implicaties van EXO1b, is het een uitstekende keuze voor S-fase specifieke rekruteringsstudies met behulp van PCNA.
Protocol
1. Kweek van van menselijke osteosarcoom afgeleide cellen (U-2 OS)
OPMERKING: U-2 OS-cellen zijn ideaal voor deze studies omdat ze een platte morfologie hebben, een grote kern en sterk hechten aan verschillende oppervlakken, waaronder glas. Andere cellijnen met vergelijkbare kenmerken kunnen ook worden gebruikt.
- Gebruik voor de teelt van U-2 OS-cellijnen McCoy's 5A-medium aangevuld met 10% foetaal runderserum (FBS) en antibiotica (100 U / ml penicilline en 100 μg / ml streptomycine). Incubeer cellen bij 37 °C in een bevochtigde atmosfeer die 5% CO2bevat. Voor microscopiestudies, bewaar celkweek in een schotel van 10 cm om voldoende celgetal te bieden.
- Wanneer cellen 90% confluentie naderen (~ 7 x 106 cellen / 10 cm schotel), splits de cellen.
- Spoel cellen met PBS om trypsineremmers in het serum weg te spoelen.
- Voeg 1 ml Trypsine-EDTA toe en zorg ervoor dat de cellaag gelijkmatig bedekt is.
- Incubeer bij 37 °C totdat de cellaag van de plaat is getild (ongeveer 6 min).
- Resuspend de trypsinized cellen in serum bevattende media om de trypsine te inactiveren en voeg 1/10e van het volume (~ 0,7 x 106 cellen) toe aan een nieuwe plaat van 10 cm met 10 ml aangevuld groeimedium.
- Voorafgaand aan de experimenten, routinematig testen cellen op mycoplasma besmetting met behulp van de Universal Mycoplasma Detection kit op aanbeveling van de fabrikant.
2. Retrovirale infectie
OPMERKING: Voor BSL-2 veiligheidsmaatregelen en tijdens het werken met recombinante virussen, raadpleegt u: NIH-richtlijnen, sectie III-D-3: Recombinante virussen in weefselkweek.
- Zaad 4 x 106 HEK293T-cellen om ~ 60% confluentie te bereiken binnen 24 uur na plating in een kweekschaal van 10 cm.
- Volg voor het kweken van HEK293T de teeltstappen van U-2 OS beschreven in 1.1-1.3 van dit protocol. Voor HEK293T vervangt u McCoy's 5A medium voor DMEM. Zorg ervoor dat u HEK293T-cellen altijd voorzichtig wast, omdat ze zich zwak hechten aan weefselkweekplaten.
- Transfect HEK293T-cellen met behulp van een op lipiden gebaseerd transfectiereagens voor virale verpakking van plasmiden.
- Combineer voor retrovirale vectoren 1,5 μg VSV-G (Addgene #8454) en 1,5 μg pUMVC (Addgene #8449) verpakkingsvectoren samen met 3 μg van de vector die het gen van belang bevat (in een retrovirale vectorbackbone met puromycineresistentie) in 250 μL Opti-MEM gereduceerde serummedia in een microcentrifugebuis. Voeg 1 μL P3000-reagens toe voor elke μg DNA die aan het Opti-MEM/DNA-mengsel wordt toegevoegd (in dit geval 6 μL) en meng voorzichtig door te tikken. Draai niet en pipetteer niet op en neer.
- Combineer in een andere microcentrifugebuis 2 μL per μg DNA (in dit geval 12 μL) transfectiereagens met 250 μL Opti-MEM gereduceerde serummedia.
- Combineer de twee mengsels (500 μL gecombineerd, draai geen vortex, meng alleen door zachtjes te tikken) en laat het 15 minuten incuberen bij kamertemperatuur.
- Voeg het mengsel voorzichtig druppelsgewijs toe aan de gezaaide HEK293T-cellen zonder de cellen los te maken. Draai de borden zachtjes rond.
- Virale infectie om stabiele cellijnen te genereren.
- Verwijder het virus dat supernatant bevat 72 uur na transfectie uit de HEK293T-cellen. Filter de oplossing zorgvuldig met een filter van 0,45 μm om celresten en losgemaakte cellen te verwijderen. Voeg eventueel 8 μg/ml polybren toe aan het virale supernatant om virale infectie te vergemakkelijken.
- Voeg virus bevattende supernatant toe aan U-2 OS-cellen met ~ 50% confluency in een schaal van 10 cm (~ 3 x 106 cellen). Zaai de U-2 OS-cellen de dag ervoor.
- Infecteer gedurende 6-16 uur voordat u het virusbevattende supernatant verwijdert en weggooit.
OPMERKING: Om de gewenste hoeveelheid overexpressie voor het gen van belang te bereiken, incubeer een reeks virale verdunningen gedurende een bepaalde tijd. Controleer de expressieniveaus van het transgen in elke nieuw vastgestelde cellijn met western blot die het vergelijkt met endogene niveaus. - Laat cellen selecteren in de aanwezigheid van geschikte antibiotica (gedurende 3-4 dagen in het geval van puromycine bij 2 μg / ml eindconcentratie) en verifieer de expressie van het fluorescerende eiwit gelabelde gen van belang onder een microscoop.
- Herhaal deze stappen om dubbel gelabelde cellijnen te genereren. In de hier gepresenteerde experimenten werd mPlum-PCNA uitgedrukt uit een retrovirale vector (pBABE) gecombineerd met EXO1B-AcGFP, ook uitgedrukt uit een retrovirale vector (pRetroQ-AcGFP1-N1).
3. Voorbereiding van cellen voor microstraling
- Platingcellen: 24 uur voor het experiment, plaat een totaal van 8,0 x 104 cellen in een volume tussen 500 μL-1 ml media (voor ongeveer 70% confluentie) op een vier goed gekamerde afdekglas met een nr. 1,5 borosilicaat glazen bodem die ideale resultaten levert voor confocale microscopie met hoge vergroting en lasermicrobestraling. Een hogere celconfluïtentie zorgt voor meer cellen gemeten in een enkel gezichtsveld (FOV); volledig samenvloeiend glaasjes zullen echter onregelmatigheden in de celcyclus introduceren.
- Beeldvormende media: Wissel een uur voor microbestraling regelmatig groeimedium uit voor FluoroBrite DMEM aangevuld met 10% FBS, 100 U/ml penicilline en 100 μg/ml streptomycine, 15 mM HEPES (pH=7,4) en 1 mM natrium-pyruvaat. Dit beeldmedium helpt de signaal-ruisverhouding te maximaliseren, waardoor zeer zwakke fluorescentie kan worden gedetecteerd. Omdat het HEPES bevat, stabiliseert het ook de pH in afwezigheid van een 5% CO2-atmosfeer.
- Pas in deze stap een eventuele aanvullende behandeling toe vóór de beeldvorming. In de hier gepresenteerde experimenten werden cellen een uur voor de beeldvorming voorbehandeld met olaparib (PARP-remmer, bij 1 μM eindconcentratie) of een voertuigcontrole (DMSO)1,8,9.
4. Het voorbereiden van de microscoop en het selecteren van S-fase cellen voor beeldvorming.
- Gebruik een confocaal systeem dat dezelfde eigenschappen heeft als het systeem dat hier wordt beschreven voor de beste resultaten. De hier gepresenteerde experimenten werden uitgevoerd met behulp van een confocale microscoop gemonteerd op een omgekeerde microscoopstandaard (zie Materiaaltabel).
OPMERKING: De microscoop die hier werd gebruikt, was uitgerust met een 50 mW 405 nm FRAP-lasermodule en een 60x 1.4 NA olieplan-apochromaatobject objectief. De confocale scankop had twee scanneropties: een galvanoscanner (voor hoge resolutie) en resonantiescanner (voor snelle beeldvorming).- Introduceer fluorescentieherstel na fotobleaching (FRAP) laser in het monster via een softwaregestuurd XY-galvano-apparaat. Gebruik een 488 nm laserlijn om AcGFP te exciteren en een 561 nm of 594 nm laserlijn om mPlum te exciteren.
OPMERKING: De volgende filtercombinatie geeft optimale resultaten: met behulp van een 560 nm langdoorlaatfilter werd emissielicht met een golflengte lager dan 560 nm door een 525/50 nm emissiefilter voor AcGFP gevoerd, terwijl emissielicht met een golflengte hoger dan 560 nm door een 595/50 nm emissiefilter voor mPlum werd gevoerd. Elke geschikte filterset (bijv. FITC/TRITC, GFP/mCherry, FITC/TxRed) die zorgt voor minimale fluorescentiedoorbloeding kan worden gebruikt.
- Introduceer fluorescentieherstel na fotobleaching (FRAP) laser in het monster via een softwaregestuurd XY-galvano-apparaat. Gebruik een 488 nm laserlijn om AcGFP te exciteren en een 561 nm of 594 nm laserlijn om mPlum te exciteren.
- Schakel de omgevingskamer en de microscoopcomponenten in.
- Schakel de verwarming (fase, objectief en omgevingskamer indien mogelijk), CO2-toevoer en de vochtigheidsregelaar ten minste 4 uur voor het begin van het experiment in om thermische evenwichtsoverbreking te garanderen voor stabiele beeldacquisitie.
- Initialiseer lichtbronnen samen met de laserlijnen ten minste 1 uur voor de overdracht van de cellen naar de microscoop.
- Selecteer S-fasecellen in een asynchrone populatie met fluorescerend getagd PCNA als marker. Volg hiervoor de onderstaande stappen.
- Zoek naar het unieke lokalisatiepatroon van het mPlum-gelabelde PCNA in S-fase waardoor identificatie van deze celcyclusfase mogelijk is. PCNA heeft een volledig homogene verdeling in de kern in G1- en G2-fasen van de celcyclus, terwijl het wordt uitgesloten van de nucleoli. In de S-fase vormt PCNA foci op de plaats van replisomen in de kern. Figuur 1 toont de verschillende patronen van PCNA-foci gedurende de S-fase, wat het mogelijk maakt om zelfs de vroege, midden- en late S-fase te differentiëren.
- Kijk door het oculair om een FOV te selecteren die voldoende S-fasecellen heeft voor micro-bestraling. Asynchrone U-2 OS-cellen hebben meestal 30-40% van hun populatie in de S-fase.
- Probeer extremen in expressieniveaus (zowel heldere als zwakke cellen) te vermijden voor zowel PCNA als het eiwit van belang (POI), in dit geval EXO1b-AcGFP, wat zou kunnen leiden tot experimentele artefacten.
- Probeer bij het vinden van een geschikte FOV te voorkomen dat het veld lange tijd wordt gescand om fotobleaching en ongewenste DNA-schade te minimaliseren.
- Stel het gewenste interessegebied (ROI) in voor microbestraling. Stel met behulp van de bijbehorende software (zie Tabel met materialen)de gewenste ROI in door eerst binaire lijnen in te voegen (stel het gewenste aantal regels en afstand in). Klik op Binairen klik vervolgens op Regel invoegen | Cirkel | Ellips om het gewenste aantal lijnen te tekenen.
- Zet deze binaire lijnen om in ROIs en zet deze ROIs uiteindelijk om in stimulatie-ROM's. Om dat te doen, klikt u eerst op ROI,vervolgens op Binair verplaatsen naar ROI,klikt u vervolgens met de rechtermuisknop op een van de ROI's en selecteert u Gebruiken als stimulatie ROI: S1. Plaats deze lijnen in de FOV om door de kern van de cellen te gaan. ROIs met een lengte van 1024 pixels die de hele FOV overspanden, werden in het hele protocol gebruikt.
5. Microbestraling voor immunofluorescentiekleuring of timelapse beeldvorming.
- Het bepalen van optimale microstralingsinstellingen.
- Maak vóór microbestraling van de cellen een afbeelding met een hogere resolutie van de FOV om PCNA-foci te identificeren voor latere analyse. In plaats van sequentieel scannen, registreert u tegelijkertijd beide gebruikte optische kanalen (groen en rood), om celbewegingen tussen scannen op de twee golflengten te voorkomen. Gebruik voor een goede resolutie van de foci ten minste 1024 x 1024 pixels / veldresolutie met 1x zoom (0,29 μm pixelgrootte op het hier gebruikte beeldvormingssysteem), met 1/8 frame / s scansnelheid (4,85 μs / pixel) met 2x gemiddelde. Zodra deze parameters zijn ingesteld in de A1 LFOV Compact GUI en de A1 LFOV Scan Area-vensters, drukt u op de knop Vastleggen om de FOV op te nemen.
OPMERKING: Het is belangrijk om tijdens experimenten dezelfde pixelgrootte te behouden om vergelijkbare resultaten te garanderen. - Om de microbestraling in te stellen, opent u het tabblad ND-stimulatie in de beeldvormingssoftware om toegang te krijgen tot het venster Tijdschema (A1 LFOV / Galvano-apparaat). Deze gebruikt de galvanoscanners om een reeks voorstimulatiebeelden te verkrijgen, te stimuleren (met behulp van de LUN-F 50 mW 405 nm FRAP-laser) en vervolgens opnieuw een reeks poststimulatiebeelden te verkrijgen met behulp van de galvanoscanners. Stel eerst drie fasen in het venster Tijdschema in. Selecteer in de kolom Acq/Stim De | Bleek | Acquisitie voor respectievelijk de drie fasen. Stel voor de bleekfase S1 in als roi.
OPMERKING: In het hier gepresenteerde experiment werden geen beelden verkregen tijdens de stimulatiefase. - Stel in het Galvano XY-vensterde belangrijkste factoren voor microstraling in: 405 nm laservermogen, verblijftijd (iteratie is standaard 1 op dit systeem). In de hier gepresenteerde experimenten werden cellen bestraald met de 405 nm FRAP-laser (50 mW aan de vezelpunt) bij 100% vermogen met een verblijftijd van 1000-3000 μs.
OPMERKING: Omdat de verblijftijd van de laser per pixel is, zolang de pixelgrootte hetzelfde blijft, zal de relatie tussen de verblijftijd en de vermogensdichtheid vergelijkbaar zijn tussen verschillende FOV's. Figuur 2A toont het gebruik van DNA damage response (DDR) pathway specifieke eiwitten (FBXL10 voor DSB's en NTHL1 voor oxidatieve basisschade) om de laservermogensinstellingen voor specifieke schade-inductie te optimaliseren. Deze stabiele cellijnen werden gegenereerd met virale infectie volgens sectie 2 van het protocol.
- Maak vóór microbestraling van de cellen een afbeelding met een hogere resolutie van de FOV om PCNA-foci te identificeren voor latere analyse. In plaats van sequentieel scannen, registreert u tegelijkertijd beide gebruikte optische kanalen (groen en rood), om celbewegingen tussen scannen op de twee golflengten te voorkomen. Gebruik voor een goede resolutie van de foci ten minste 1024 x 1024 pixels / veldresolutie met 1x zoom (0,29 μm pixelgrootte op het hier gebruikte beeldvormingssysteem), met 1/8 frame / s scansnelheid (4,85 μs / pixel) met 2x gemiddelde. Zodra deze parameters zijn ingesteld in de A1 LFOV Compact GUI en de A1 LFOV Scan Area-vensters, drukt u op de knop Vastleggen om de FOV op te nemen.
- Timelapse beeldvorming.
- Stel timelapse-beeldvorming in voor het gewenste tijdvenster en intervallen met behulp van het tijdschema, de A1 LFOV Compact GUI en de vensters A1 LFOV Scan Area. In de hier gepresenteerde experimenten werd de rekrutering van EXO1b en PCNA gedurende 12 minuten in beeld gegeven, waarbij de FOV elke 5 seconden werd gescand, op 1024 x 1024 pixels / veld, met behulp van 1x zoom (resulterend in 0,29 μm pixelgrootte op het beeldvormingssysteem dat hier wordt gebruikt) met een scansnelheid van 0,35 frame / s (1,45 μs / pixel) zonder middeling om fotobleek te verminderen.
- Optimaliseer het laservermogen % , versterking en offset instellingen om foto-bleken tijdens de beeldvorming in het A1 LFOV Compact GUI-venster te verminderen. Als men zowel POI als PCNA wil meten, gebruik dan gelijktijdig scannen in plaats van sequentieel scannen om celbewegingen tussen het scannen van het veld voor de twee afzonderlijke fluoroforen te voorkomen.
- Het beeldvormingssysteem werd gebruikt met de volgende instellingen. Voor de 488 nm laserlijn (20 mW): 7% laservermogen, versterking: 45 (GaAsP detector) met en offset van 2, voor de 561 nm laserlijn (20 mW): 4% laservermogen, gain 40 (GaAsP detector) met en offset van 2.
- Afhankelijk van de kinetiek van het eiwit, verleng of verkort het interval tussen beelden of de duur van het totale tijdsverloop. Stel in het venster Tijdsplanning het gewenste interval en de gewenste duur in voor de rij Acquisitie van de derde fase.
- Druk nu op Run om de microbestraling en de daaropvolgende time-lapse-beeldvorming uit te voeren.
- Sla aan het einde van de time-lapse-beeldvorming de stimulatie-ROM's op als afzonderlijke afbeeldingen, wat een nuttig hulpmiddel zal zijn om de coördinaten van microbestraling te identificeren in alle downstream-software die voor analyse wordt gebruikt.
- Immunofluorescentiekleuring.
OPMERKING: Stap 5.1.3 en figuur 2A tonen het gebruik van bekende DNA-reparatie-eiwitten om de soorten DNA-laesies te beoordelen die door microbestraling worden geïntroduceerd. Bepaalde DNA-laesies kunnen ook worden gedetecteerd met behulp van specifieke antilichamen na het fixeren van de cellen. Het is ook mogelijk om de rekrutering van de POI te detecteren door antilichaamdetectie van het endogene eiwit. De visualisatie van γH2A.X om te controleren op DSB's wordt hieronder gedemonstreerd(Figuur 2B). Figuur 3 toont de consistentie van PCNA-lokalisatie en -rekrutering gedurende de hele celcyclus voor zowel endogene als exogene gelabelde PCNA.- Neem na stap 5.1.3 slechts één beeld na microbestraling om de juiste FRAP-gebeurtenis te garanderen op basis van de rekrutering van mPlum-PCNA. Let op de exacte coördinaten van de FOV om het veld later na de immunofluorescente labeling te vinden.
- Haal de celkweek uit de microscoop en incubeer cellen bij 37 °C in een bevochtigde atmosfeer met 5% CO2 gedurende 5-10 minuten.
OPMERKING: Paraformaldehyde (PFA) is giftig en het werk moet worden uitgevoerd in een goed geventileerde ruimte of een zuurkast. Alle daaropvolgende wassing en incubatie zal worden gedaan met 0,5 ml volumes in de 4 putkamerschuif. Was na de incubatietijd de cellen met 0,5 ml PBS (137 mM NaCl, 2,7 mM KCl, 8 mM Na2HPO4en 2 mM KH2PO4) en fixeer met 0,5 ml van 4% PFA in PBS gedurende 10 minuten bij kamertemperatuur (RT). - Was de cellen eenmaal met PBS en was ze vervolgens met 50 mM NH4Cl om resterende PFA te dempen.
- Permeabiliseer de cellen gedurende 15 minuten bij RT met 0,1% Triton X-100 in PBS.
- Blokkeer de monsters gedurende 1 uur met blokkeringsbuffer (5% FBS, 3% BSA, 0,05% Triton X-100 in PBS).
- Verwijder de blokkerende oplossing en voeg het verdunde primaire antilichaam (anti-γH2A.X, 1:2000) toe in de blokkeringsbuffer gedurende 1 uur bij RT.
- Was de putjes met blokkeerbuffer 3 x 10 min.
- Voeg verdund secundair antilichaam (anti-muis Alexa 488 Plus-conjugaat, 1:2000) toe in de blokkeringsbuffer gedurende 1 uur bij RT.
- Was de putjes met blokkeerbuffer 3 x 10 min.
- Counterstain de kern met 1 μg/ml DAPI-oplossing in PBS gedurende 15 minuten.
- Was de cellen eenmaal met PBS. De beeldvorming kan rechtstreeks worden uitgevoerd in PBS of een PBS-oplossing met antifadereagentia (bijv. AFR3) om fotobleeken te verminderen.
6. Rekruteringsanalyse
OPMERKING: Figuur 4A toont representatieve beelden van Exo1b- en PCNA-rekrutering in aanwezigheid van DMSO of olaparib. Figuur 4B toont een representatief beeld voor data-analyse. De gemiddelde fluorescentiewaarden werden berekend door de gemiddelde AcGFP-intensiteiten te meten met behulp van een rechthoek langs de laserbaan gemarkeerd door de mPlum-PCNA (A, gele rechthoeken) over verschillende tijdspunten met behulp van Fiji. PCNA kan dienen als een interne controle om succesvolle bestraling langs de ROI-coördinaten te markeren. Evenzo werden ook gemiddelde AcGFP-fluorescentiewaarden berekend voor onbeschadigde gebieden van de kern (B, blauwe rechthoeken). De intensiteit van het achtergrondsignaal werd gemeten in onbevolkte gebieden (C, rode rechthoeken) en werd afgetrokken van de gemiddelde fluorescerende waarden(figuur A en B). Zo werd de relatieve gemiddelde fluorescerende eenheid (RFU) voor elk gegevensverzamelingspunt berekend door de vergelijking RFU = (A − C)/ (B − C)8,9. De resulterende RFU-waarden van het microbestraalde gebied worden genormaliseerd tot de RFU-waarden voorafgaand aan microbestraling.
- Voor het definiëren van het gebied A van de microbestraalde locatie, sluit nucleolaire gebieden, replicatiehaarden en onregelmatige nucleaire gebieden van de cel uit van de meting. Houd de shift-toets ingedrukt tussen het tekenen van twee RO's in Fiji om twee afzonderlijke regio's als één te groeperen.
OPMERKING: Eiwitrekrutering zal variëren tussen verschillende genen en bestralingsomstandigheden; de grootte van regio A moet dus individueel worden bepaald. Zodra de pixelbreedte van gebied A is bepaald, moet deze constant blijven voor vergelijkende rekruteringen. In de hier gepresenteerde experimenten werden rechthoeken met een breedte van 7 pixels gebruikt. - Sluit cellen die tijdens de duur van de opgenomen video's zijn verplaatst, uit van analyse. Om zeer mobiele cellen op te nemen, moet de beschreven analyse frame voor frame worden uitgevoerd.
- Om het wervingsprofiel te visualiseren, plot u de genormaliseerde RFU-waarden tegen de tijd met behulp van statistische software.
- Bereken het verschil op een aangegeven tijdstip tussen DMSO en olaparib (n=31) behandeling met behulp van een Mann-Whitney test.
Representative Results
Cellen pakken elk type DNA-laesie aan op een specifieke manier die ook afhangt van de fase van de celcyclus waarin ze zich bevinden. Na microbestraling worden bijvoorbeeld dubbelstrengs breuken (DSB) verwerkt door niet-homologe eindvoeging (NHEJ) of HR, afhankelijk van de celcyclusfase. Nucleasen die het meest uitgebreid werken tijdens de S- en G2-fasen van de celcyclus creëren DNA-overhangen die cruciaal zijn voor een goede HR. Om de evaluatie van cellen in de S-fase te bevorderen, werd PCNA gebruikt als een eenkleurige celcyclusmarker. Figuur 1A toont het lokalisatieprofiel van mPlum-PCNA tijdens de progressie van de celcyclus. PCNA heeft een volledig homogene verdeling in de kern in de G1- en G2-fase (terwijl het ook meestal wordt uitgesloten van de nucleoli). In de S-fase lokaliseert PCNA naar plaatsen van DNA-replicatie, die kunnen worden gevisualiseerd als heldere vlekken in de kern. In vroege S-fase cellen zijn de vlekken relatief klein en gelijk verdeeld over de kern van de cel. In het midden van de S-fase worden de vlekken wazig en lokaliseren ze meer naar de omtrek van de kern en de nucleoli. In de late S-fase nemen de vlekken in aantal af, maar worden ze steeds groter naarmate PCNA zich concentreert op late replicatielocaties(figuur 1B). Belangrijk is dat exogene PCNA-expressie van de pBABE-vectorbackbone minder was dan de endogene niveaus, maar voldoende was voor detectie door microscopie die potentiële artefacten in celcyclusprogressie en DDR minimaliseert. Figuur 1C toont de mate van PCNA-overexpressie in vergelijking met endogene niveaus. Houd er rekening mee dat de band die overeenkomt met mPlum-PCNA langzamer migreert vanwege het grotere formaat.
We wilden DSB's introduceren tijdens microbestraling om de PARP1/2-afhankelijke rekrutering van EXO1b voor deze laesies in de S-fase te onderzoeken. Figuur 2A laat zien dat lage doses energie (1000 μs verblijftijd) niet leiden tot de rekrutering van EGFP-FBXL10, een DSB-responder (onderdeel van het FRUCC-complex 8), terwijl het voldoende was om de rekrutering van NTHL1-mCherry, een base excision repair (BER) pathway-eiwit, te induceren naar plaatsen van oxidatieve DNA-schade10,11,12. Bij 3000 μs verblijftijd, zowel EGFP-FBXL10 als NTHL1-mCherry rekruteren, wat een laseroutput aantoont die zowel oxidatieve laesies als DSB's genereert. Figuur 2B versterkt deze resultaten en toont immunofluorescentiekleuring tegen γH2A.X (DSB-marker), die duidelijk duidelijker is bij het gebruik van hogere energiedoses. PCNA dient als zowel een celcyclusmarker als een marker voor succesvolle microbestraling, omdat het adequaat rekruteert met beide laserbewoningstijdinstellingen. Belangrijk is dat zowel exogene als /of endogene fluorescerende eiwit-gelabelde PCNA kan worden gebruikt voor deze reporterfunctie, omdat ze zich op dezelfde manier gedragen(figuur 3). Endogeen getagde PCNA werd ontworpen door mRuby in frame met het eerste exon in één allel van de PCNA locus13 in te brengen (de cellijn was een soort geschenk van Jörg Mansfeld).
Figuur 4A en figuur 4C tonen de rekrutering van AcGFP-gelabelde EXO1b in S-fasecellen. EXO1b bereikt ongeveer 1 minuut het maximale accumulatieniveau op microbestralingsplaatsen en begint daarna langzaam los te weken van de DNA-laesies. Verrijkingen op microstralingsplaatsen worden aangegeven door een > 1 relatieve fluorescentie-eenheid op de grafiek. In aanwezigheid van olaparib is de accumulatie van EXO1b op de laserstreep na 1 minuut aanzienlijk minder in vergelijking met de voertuigbesturing. Deze resultaten komen in overeenstemming met de literatuur6,7. Figuur 4B toont representatieve regio's voor kwantificering (gebieden A, B en C) zoals beschreven in punt 6 van het protocol. Figuur 4D toont de vergelijkbare expressieniveaus van endogene EXO1b en exogene EXO1b-AcGFP in cellen die worden gebruikt voor microstraling.
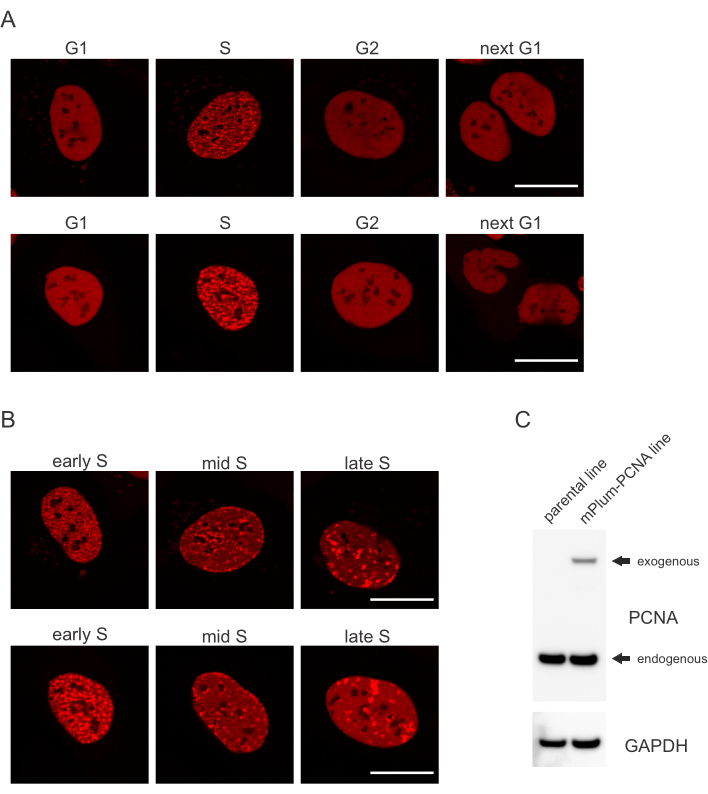
Figuur 1: Lokalisatiepatroon van PCNA. (A) Afbeeldingen tonen het lokalisatiepatroon van stabiel geïntegreerd, exogene PCNA gedurende de celcyclus in U-2 OS-cellen. (B) Afbeeldingen tonen PCNA-focipatronen in verschillende stadia van de S-fase (vroeg, midden en laat) in U-2 OS-cellen. (C)Western blot met endogene en exogene niveaus van PCNA in de U-2 OS-cellen die worden gebruikt voor beeldvorming. Schaalbalk vertegenwoordigt 20 μm. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 2: Inductie van DSB's door geoptimaliseerd laservermogen. (A) Laserinstellingen kunnen worden geoptimaliseerd om verschillende vormen van DNA-schade te induceren. U-2 OS-cellen die zowel EGFP-FBXL10 als NTHL1-mCherry stabiel tot expressie brengen, werden gebruikt om respectievelijk DSB's en plaatsen van oxidatieve laesies te identificeren. Microbestraling met een 405 nm laserlijn werd uitgevoerd op asynchrone U-2 OS-cellen met een verblijftijd van 1000 μs of 3000 μs. Schaalbalk vertegenwoordigt 20 μm. (B) Immunofluorescente kleuring tegen γH2A.X werd gedaan op menselijke retinale pigmentepitheelcellen (hTERT RPE-1) met mRuby-tagged endogene PCNA. Cellen werden gefixeerd en verwerkt 5 minuten na micro-bestraling met een verblijftijd van 1000 μs of 3000 μs. Schaalbalk vertegenwoordigt 20 μm. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 3: Vergelijkbare rekrutering van endogene mRuby-PCNA en exogene mPlum-PCNA naar microbestralingsplaatsen bij 1000 μs of 3000 μs lasertijd. Zowel endogene als exogene gelabelde PCNA vormen replicatiehaarden tijdens de S-fase. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 4: PARP1/2-afhankelijke rekrutering van EXO1b in S-fase. U-2 OS-cellen die stabiel EXO1b-AcGFP en mPlum-PCNA tot expressie brengen, werden microbestraald met 405 nm FRAP-laserlijn met behulp van 3000 μs verblijftijd. A)Representatieve beelden van microbestraalde cellen op de aangegeven tijdstippen na voorbehandeling met mediumcontrole (DMSO) of olaparib (1 μM). Schaalbalk vertegenwoordigt 20 μm. (B) Representatieve afbeeldingen van gedefinieerde gebieden van A-, B- en C-gebieden voor de wervingsanalyse. Schaalbalk vertegenwoordigt 20 μm. (C) DNA-schade rekruteringsdynamiek werd vastgelegd door live celbeeldvorming. Relatief gemiddelde fluorescentiewaarden en beelden werden elke 5 s gedurende 12 minuten verkregen. Voor elke aandoening werden ≥30 cellen geëvalueerd. Gemiddelde relatieve fluorescentiewaarden (ononderbroken zwarte lijnen) en standaardfout (bereik gevisualiseerd door een gearceerd gebied) werden uitgezet tegen de tijd. Stippellijn toont rekruteringswaarden op 1 minuut na micro-bestraling. Het verschil tussen de behandeling met DMSO (n=32) en olaparib (n=31) werd berekend met behulp van een Mann-Whitney-test. Asterix duidt op p<0,0001. (D) Western blot vergelijkt de expressieniveaus van endogene EXO1b en exogene EXO1b-AcGFP in cellen die worden gebruikt voor microbestraling. Klik hier om een grotere versie van deze figuur te bekijken.
Discussion
Kritieke stappen en mogelijke probleemoplossing/wijzigingen in het protocol
Een goed weefselkweekvat voor microbestraling is van cruciaal belang voor succes. De meeste beeldvormingssystemen met hoge resolutie zijn geoptimaliseerd voor een dikte van 0,17 mm afdekglas. Het gebruik van beeldvormingskamers met een hogere of lagere dikte of kamers gemaakt van plastic polymeren (niet geoptimaliseerd voor 405 nm-beeldvorming), kan de beeldkwaliteit aanzienlijk verminderen. Wanneer u glazen oppervlakken gebruikt, moet u ervoor zorgen dat ze weefselkweek zijn behandeld om de celadhesie te verbeteren. Als ze niet met weefselkweek worden behandeld, moeten deze kamers bijvoorbeeld worden bedekt met poly-D-lysine voordat de cellen worden gezaaid. Bij het platen van cellen in het kamerdekglas is een ideale celdichtheid van het grootste belang om onregelmatigheden in de celcyclus en extra stress voor de cellen te voorkomen. Een goede thermische evenwichtsverdeling van de microscoopcomponenten voorafgaand aan het experimenteren om een stabiele temperatuur te behouden, is cruciaal voor zowel het behoud van de focus gedurende de time-lapse-beeldvorming als is ook noodzakelijk om een homogene DDR in de tijd en monsters te garanderen.
Het is van cruciaal belang dat cellen zich in een gezonde toestand bevinden voorafgaand aan microbestraling om artefactuele gegevens te verminderen. Als cellen een onregelmatige morfologie hebben na infectie / selectie, laat cellen dan door meerdere passages gaan totdat de morfologie weer normaal wordt. Zorg er altijd voor dat de gebruikte cellijnen vrij zijn van mycoplasmabesmetting. Onder de vele nadelige effecten van mycoplasma-infectie, veroorzaakt het ook DNA-schade aan de gastheercellen en kan het hun DDR-routesbeïnvloeden 14,15. De meest gevoelige manier om mycoplasma in de celkweek te detecteren is via PCR (versus detectie met DAPI of Hoechst).
Optimale overexpressie van het reparatie-eiwit van belang moet vergelijkbaar zijn met endogene niveaus, maar hoog genoeg voor detectie. De promotor die wordt gebruikt op de virale vectoren, de virale titer tijdens infectie en de lengte van de infectietijd kunnen allemaal worden aangepast voor ideale expressieniveaus. Voor consistente resultaten kunt u individuele celklonen isoleren om homogene expressieniveaus en normale celmorfologie te garanderen. Het wordt aanbevolen om vectorconstructen te gebruiken die gelabelde PCNA niet overexpressie op hogere dan endogene niveaus voor een goede celcyclus en DNA-reparatiemarkerfunctie. Zelfs lage niveaus van PCNA-overexpressie zijn voldoende om S-fasecellen te onderscheiden. Retrovirale pBABE-vectoren zijn met succes voor dit doel gebruikt (Addgene #1764, #1765, #1766, #1767). PCNA kan worden getagd met monomere rode(bijv. mPlum, mCherry, mRuby, enz.) of monomere groene fluorescerende eiwitten (bijv. mEGFP, AcGFP, mWasabi, mNeonGreen, mEmerald, enz.) die vervolgens kunnen worden gecombineerd met een afwisselend getagde POI. Het overexpresseren van een fluorescerend getagde POI heeft enkele beperkingen en overwegingen. Fluorescerende tags kunnen de normale eiwitfunctie en lokalisatie verstoren. Er moet dus rekening worden gehouden met de locatie van de tag (N- of C-terminal). Gebruik altijd monomere fluorescerende eiwitten, omdat oligomerisatie van niet-monomere varianten de functie van de POI kan beïnvloeden.
De laserinstellingen moeten voor elk beeldvormingssysteem worden bepaald, omdat veel componenten van het optische pad van invloed zijn op het werkelijke vermogen dat in de cellen wordt geleverd. Lasermicrobestraling kan verschillende soorten DNA-laesies veroorzaken, afhankelijk van de excitatiegolflengte, het uitgangsvermogen van de FRAP-laser en of er voorsensibiliserende middelen (zoals Bromodeoxyuridine of Hoechst) zijn gebruikt. 405 nm lasers kunnen oxidatieve DNA-schade veroorzaken, enkel- en dubbelstrengs breuken16,17. Door hogere laseruitvoerinstellingen te gebruiken, neemt het aantal DSB's toe. In dit protocol werden pre-sensibilisatiemethoden niet gebruikt, maar deze technieken worden sterk behandeld in de literatuur en opnieuw afgetopt in de onderstaande discussie. Naar onze mening is de beste manier om te testen of de gewenste laesie wordt gegenereerd, door te testen op de rekrutering van bekende DNA-schaderoutespecifieke genen. Rekrutering van NTHL1 of OGG1, componenten van de BER-route, suggereert de inductie van geoxideerde DNA-basen10,11,17,18,19,terwijl FBXL10 of XRCC5 de aanwezigheid van DSBs8,20,21aangeven. Rekrutering van XRCC1 kan zowel de aanwezigheid van geoxideerde DNA-basen als enkelstrengsbreuken (SSB)aangeven 22,23. XPC (d.w.z. RAD4) is een goede indicator van NER die de omvangrijke DNA-adducten verwijdert die worden gegenereerd door ultraviolet licht (UV)17,24. Omdat het rekruteren van exogene eiwitten bepaalde onregelmatigheden kan introduceren, kan immunofluorescente kleuring van endogene DNA-reparatie-eiwitten of markers (zoals γH2A.X voor dubbelstrengsbreuken) de aanwezigheid van specifieke DNA-laesies bevestigen. Als alternatief kunnen ook antilichamen tegen specifieke soorten DNA-laesies worden gebruikt. Om het geleverde laservermogen aan te passen, kunnen zowel de verblijftijd als het laservermogen worden gewijzigd.
Met behulp van wiskundige modellering kan een gedetailleerde kinetische analyse worden uitgevoerd die waardevolle inzichten kan bieden in de rekruteringseigenschappen van de POI (bijv. Bijdrage van meerdere DNA-bindingsdomeinen, gevoeligheid voor verschillende signaleringsgebeurtenissen, enz.). Geautomatiseerde wervingsevaluatie en celtracking kunnen worden gecombineerd om robuuste workflows 1,25te creëren.
Voordelen en beperkingen van DNA-presensibilisatie
Presensibilisatie van DNA voorafgaand aan micro-bestraling is een veelgebruikt hulpmiddel voor DNA-reparatie eiwitrekrutering16,17. Sensibiliserend DNA voorafgaand aan microbestraling maakt het gevoeliger voor DSB's. De twee meest voorkomende methoden voor DNA-voorsensibilisatie zijn voorbehandeling van cellen met Bromodeoxyuridine (BrdU) of Hoechst-kleurstof. Voor systemen die niet in staat zijn tot microbestraling met hoge laservermogens, kunnen deze methoden nodig zijn voor het induceren van DNA-laesies zoals DSB's. Bovendien, bij afwezigheid van een doorgegeven lichtdetector of een fluorescerend signaal dat de celkern benadrukt (bijvoorbeeld bij het bestuderen van de rekrutering van niet-gecodeerde endogene DNA-reparatie-eiwitten), fungeert Hoechst als zowel een presensibiliserend hulpmiddel als een fluorescerende nucleaire kleuring. DNA-presensibilisatie kan echter aanzienlijke complicaties met zich meebrengt. BrdU (gebruikt bij een eindconcentratie van 10 μM) moet 24 uur (of tijdsequivalent aan een volledige celcyclus in de gebruikte cellijn) worden toegevoegd om correct in het DNA te worden opgenomen en kan celcyclusinterferentie veroorzaken26. Hoechst 33342 (gebruikt bij een eindconcentratie van 1 μg/ml) is cytotoxisch na lange incubatieperioden, maar heeft voldoende tijd nodig om de kern met de kleurstof te verzadigen. Daarom mag het alleen 15-20 minuten voorafgaand aan microbestraling worden aangebracht; anders zijn de wervingsgegevens niet consistent. De cellen die op deze manier worden gekleurd, kunnen niet langer dan een paar uur in cultuur worden gehouden27,28. Zorg ervoor dat u geen Hoechst 33358 gebruikt, die niet zo celdoorlatend is als de Hoechst 33342 kleurstof. Pre-sensibilisatie kan ook onnodige variatie tussen experimenten introduceren en maakt het experiment nog gevoeliger voor verschillen in celdichtheid (omdat dit de hoeveelheid opgenomen kleurstof / cel zal beïnvloeden).
Voordelen en beperkingen van confocale microscopie
De beeldvormingssnelheid van confocale microscopie kan beperkend zijn in vergelijking met widefield-microscopie. Een confocale microscoop uitgerust met een resonerende scanner kan echter de beeldvormingssnelheid enorm verbeteren (ten koste van de resolutie) en in de buurt komen van de snelheden van spinning-disk microscopie. Drie functies maken het A1R HD25 confocale systeem een uitstekende keuze voor het hier gepresenteerde protocol. Ten eerste maakt de 25 mm FOV van het systeem het mogelijk om tussen de 15-20 cellen in een enkel gescand veld (versus 5-10 cellen in reguliere opstellingen) in beeld te brengen, waardoor het aantal acquisities dat nodig is om voldoende cellen te krijgen voor statistische analyse wordt beperkt. Ten tweede maken de FRAP-module en twee scankoppen het mogelijk om de cellen tegelijkertijd in beeld te brengen en te micro-bestralen, niet alleen sequentieel. Ten slotte biedt de flexibiliteit van zowel de resonantie- als galvanoscanners de mogelijkheid om gemakkelijk te schakelen tussen beeldvorming met een hoge temporele resolutie met een uitzonderlijke snelheid die het doven van fluoroforen minimaliseert, en beeldvorming met een hoge ruimtelijke resolutie die langzamere scansnelheden gebruikt om beelden te produceren met een hogere signaal-ruisverhouding. Hoewel het gebruikte systeem de bovengenoemde flexibiliteit mogelijk maakte, om te lijken op meer algemeen beschikbare confocale microscoopconfiguraties, werd alleen de galvanoscanner gebruikt in de gepresenteerde experimenten (voor zowel microbestraling als daaropvolgende beeldvorming).
Voordelen en beperkingen van microbestraling
Hoewel microstraling een ongeëvenaarde ruimtelijke en temporele resolutie biedt, is het niet zonder beperkingen. DNA-schade door lasermicrostraling is sterk geclusterd naar specifieke delen van de kern in vergelijking met natuurlijk voorkomende schadelijke stoffen. De chromatinerespons als gevolg van microstraling kan dus verschillen van homogeen verdeelde schade. Bovendien is microbestraling tijdrovend en kan deze slechts op enkele tientallen cellen worden uitgevoerd, terwijl grote populatiegebaseerde biochemische methoden (chromatinefractionering, immunoprecipitatie, ChIP) een verhoogde robuustheid kunnen bieden door duizenden cellen tegelijk te bestuderen. Het verifiëren van waarnemingen door microbestraling met traditionele biochemische technieken is een effectieve strategie voor betrouwbare conclusies. Hoewel gelijktijdige microbestraling van veel cellen in een bepaalde FOV mogelijk is, heeft het beeldvormingssysteem meer tijd nodig om de taak uit te voeren. Daarom beperkt het meten van de dynamiek van eiwitten die zeer snel rekruteren voor DNA-laesies het aantal mogelijke ROIs voor microbestraling dat tegelijkertijd wordt gebruikt. Op het beeldvormingssysteem dat voor dit protocol wordt gebruikt, duurt de microbestraling van een enkele ROI van 1024 pixels lang 1032 ms met een verblijftijd van 1000 μs en 3088 ms met een verblijftijd van 3000 μs om te voltooien. Het gebruik van meerdere lijnen van ROI's zal de tijd die nodig is om micro-bestraling te voltooien aanzienlijk verlengen (bijv. 7 x 1024 pixel lange ROI duurt 14402 ms met een verblijftijd van 1000 μs en 21598 ms met een verblijftijd van 3000 μs). Deze tijd gaat verloren door beeldverwerving en moet in overweging worden genomen. Gebruik bij het in beeld brengen van snelle rekruteringsgebeurtenissen de kortst mogelijke ROI en micro-bestraling slechts één cel tegelijk.
Voordelen en beperkingen ten opzichte van synchronisatiemethoden
Voor celcyclusspecifieke studies omvatten de bestaande methoden ofwel de synchronisatie van cellen in specifieke celcyclusfasen of het gebruik van fluorescerende reporters om de specifieke celcyclusfase van de cel te identificeren. Elk van deze methoden biedt echter zijn eigen uitdagingen en beperkingen.
Het FUCCI-systeem3 (gebaseerd op fluorescerend eiwit met gelabelde afgeknotte vormen van CDT1 en Geminin) is een bijzonder nuttig hulpmiddel voor celcyclusstudies, maar heeft beperkingen als het gaat om het onderscheiden tussen S- en G2-fasen van de celcyclus. Geminin niveaus zijn al hoog vanaf het midden van de S-fase en blijven hoog tot de M-fase, waardoor deze fasen moeilijk te scheiden zijn. Het gebruik van het FUCCI-systeem betekent ook dat twee optische kanalen van de microscoop niet kunnen worden gebruikt voor het afbeelden van de POI.
Niet-kankercellijnen kunnen worden gesynchroniseerd met G0 door de verwijdering van groeifactoren in het serum (serumuithongering) waardoor weinig of geen DNA-schade aan de cellen ontstaat. De meeste kankercellijnen zullen echter gedeeltelijk door de celcyclus blijven gaan, zelfs zonder voldoende hoeveelheden serum in hun media. Bovendien beginnen cellen gedeeltelijk de synchronisatie te verliezen tegen het einde van de G1, vroege S-fase. Naast serumgebrek zijn er tal van chemische methoden om celcyclussynchronisatie te bereiken. Hydroxyurea, aphidicolin en thymidine-blokken zijn methoden om DNA-replicatie te stoppen om cellen te synchroniseren in de vroege S-fase. Hoewel deze methoden goedkoop en eenvoudig zijn, introduceren ze replicatiestress die resulteert in DNA-schade. Van deze DNA-replicatieremmers is aangetoond dat ze de fosforylering van H2A induceren. X, een bekende marker van DSBs2,29. De methode van het gebruik van tagged-PCNA als marker voor S-fase cellen vermindert het potentieel voor artefacten veroorzaakt door chemische synchronisatie en kan worden toegepast op een breed scala aan cellijnen in vergelijking met serumuithongering.
Conclusie
DNA-schade is een drijvende kracht voor genetische ziekten waarbij mutagene laesies kunnen leiden tot de kwaadaardige transformatie van cellen. Het richten op de DNA-synthesemachinerie is een fundamentele therapeutische strategie bij de behandeling van hyperproliferatieve ziekten zoals kanker. Om deze ziekten gerichter te behandelen, hebben we een beter begrip nodig van de eiwitten die DNA-laesies repareren. Het hier beschreven protocol helpt op microbestraling gebaseerde studies in de S-fase door de uitdagingen van traditionele synchronisatiemethoden te minimaliseren om mogelijke artefacten te verminderen en de reproduceerbaarheid van de experimenten te vergroten.
Disclosures
De auteurs stellen dat de publicatie van het gepresenteerde werk werd gesponsord door Nikon Corporation. De auteurs verklaren dat er geen tegenstrijdige belangen bestaan.
Acknowledgments
De auteurs bedanken M. Pagano voor zijn voortdurende steun, evenals D. Simoneschi, A. Marzio en G. Tang voor hun kritische beoordeling van het manuscript. B. Miwatani-Minter bedankt R. Miwatani en B. Minter voor hun voortdurende steun. G. Rona bedankt K. Ronane Jurasz en G. Rona voor hun voortdurende steun.
Materials
| Name | Company | Catalog Number | Comments |
| Ammonium chloride | Sigma-Aldrich | A9434-500G | For quenching formaldehyde |
| Anti-EXO1 Rabbit Polyclonal Antibody | Proteintech | 16253-1-AP | primary antibody |
| Anti-phospho-Histone H2A.X (Ser139) Antibody, clone JBW301 | Millipore | 05-636 | primary antibody |
| Bovine Serum Albumin | Sigma-Aldrich | 3117332001 | BSA for blocking |
| BrdU (5-Bromo-2'-deoxyuridine) | Merck | 19-160 | pre-sensitizing agent |
| Citifluor™ Mountant Solution AFR3 | Electron Microscopy Sciences | 17973-10 | antifade containing PBS solution for imaging |
| DAPI | Sigma-Aldrich | D9542-1MG | nucleic acid stain |
| DMEM Medium | Thermo Fisher Scientific | 10569010 | Cell culture medium for HEK293T cells |
| DMSO | Sigma-Aldrich | D2650-100ML | Vehichle control and dissolution solvent |
| EGFP-FBXL10 | Addgene | #126542 | viral expression vector for EGFP-FBXL10 |
| EXO1b-AcGFP (in pRetroQ) | custom cloning | na | EXO1b cDNA was cloned in the NheI, BamHI sites of pRetroQ-AcGFP1-N1 vector. |
| Fetal Bovine Serum | Gibco | 16140071 | Media supplement |
| FluoroBrite DMEM | Thermo Fisher Scientific | A1896701 | Phenol red free medium for microscopy |
| Goat anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 488 | Thermo Fisher Scientific | A32723 | secondary antibody |
| HEK293T cells | ATCC | ATCC CRL-3216 | Cell line for viral packaging |
| HEPES | Sigma-Aldrich | H0887-100ML | Buffering agent to supplement live cell imaging medium |
| Hoechst 33342 | Thermo Fisher Scientific | H3570 | pre-sensitizing agent |
| Lipofectamine 3000 | Thermo Fisher Scientific | L3000015 | Transfection reagent |
| McCoy’s 5A (Modified) Medium | Life Technologies | 16600-108 | Cell culture medium for U-2 OS cells |
| mCherry-PCNA | Addgene | #55117 | non-viral PCNA construct suitable for cell cycle marker |
| mPlum-PCNA | Addgene | #55994 | non-viral PCNA construct suitable for cell cycle marker |
| mPlum-PCNA (in pBABE) | custom cloning | na | mPlum-PCNA cDNA was cloned from Addgene #55994 in the BamHI, SalI sites of pBABE (puro) |
| Nikon A1R-HD25 Confocal Scanhead and Controller | Nikon | na | confocal imaging system |
| Nikon LUN4 laser unit | Nikon | na | excitation system |
| Nikon LUN-F 50 mW 405 nm FRAP laser unit | Nikon | na | FRAP laser unit |
| Nikon NIS Elements Confocal Controller Software | Nikon | na | Confocal controlling software |
| Nikon Ti2-E Inverted Microscope | Nikon | na | inverted epifluorescent microscope base |
| Nikon Ti2-LAPP Modular Illumination System | Nikon | na | illumination system |
| NTHL1-mCherry (in pRetroQ) | custom cloning | na | NTHL1 cDNA was cloned in the NheI, SalI sites of pRetroQ-mCherry-N1 vector. |
| Nunc Lab-Tek II Chambered Coverglass (4 well) | Thermo Fisher Scientific | 155382PK | Live cell microscopy cell culture chamber |
| Olaparib | Selleck Chemicals | S1060 | PARP inhibitor |
| Opti-MEM reduced serum media | Thermo Fisher Scientific | 31985062 | Dilution medium for transient transfection |
| Paraformaldehyde aqueous solution (32%) | Thermo Fisher Scientific | 50-980-494 | Fixative |
| pBABE (hygro) | Addgene | #1765 | retroviral expression vector (for low expression levels) |
| pBABE (neo) | Addgene | #1767 | retroviral expression vector (for low expression levels) |
| pBABE (puro) | Addgene | #1764 | retroviral expression vector (for low expression levels) |
| pBABE (zeo) | Addgene | #1766 | retroviral expression vector (for low expression levels) |
| PCNA Antibody (PC10) | Santa Cruz | sc-56 | primary antibody |
| Penicillin-Streptomycin-Glutamine (100x) | Gibco | 10378016 | Media supplement |
| polybrene | Sigma-Aldrich | TR-1003 | Increase viral infection efficiency |
| pRetroQ-AcGFP-C1 | Takara | 632506 | retroviral expression vector |
| pRetroQ-AcGFP-N1 | Takara | 632505 | retroviral expression vector |
| pRetroQ-mCherry-C1 | Takara | 632567 | retroviral expression vector |
| pRetroQ-mCherry-N1 | Takara | 632568 | retroviral expression vector |
| pUMVC | Addgene | #8449 | Viral packaging vector |
| Sodium-pyruvate | Thermo Fisher Scientific | 11360070 | Supplement for live cell imaging medium |
| Triton X-100 aqueous solution (10%) | Sigma-Aldrich | 11332481001 | Dilute in PBS for cell permeabilization buffer |
| Trypsin-EDTA Solution 10X | Sigma-Aldrich | 59418C-100ML | Dilute in PBS to split cells |
| U-2 OS Cells | ATCC | HTB-96 | Optimal cell line for microscopy experiments |
| Universal Mycoplasma Detection Kit | ATCC | 30-1012K | PCR based Mycoplasma detection kit |
| VSV-G | Addgene | #8454 | Viral protein envelope vector |
References
- Aleksandrov, R., et al.
- Darzynkiewicz, Z., Halicka, H. D., Zhao, H., Podhorecka, M. Cell synchronization by inhibitors of DNA replication induces replication stress and DNA damage response: Analysis by flow cytometry. Methods in Molecular Biology. 761, 85-96 (2011).
- Sakaue-Sawano, A., et al. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell. 132 (3), 487-498 (2008).
- Herce, H. D., Rajan, M., Lattig-Tunnemann, G., Fillies, M., Cardoso, M. C. A novel cell permeable DNA replication and repair marker. Nucleus. 5 (6), 590-600 (2014).
- Keijzers, G., et al. Human exonuclease 1 (EXO1) regulatory functions in dna replication with putative roles in cancer. International Journal of Molecular Sciences. 20 (1), (2018).
- Cheruiyot, A., et al. Poly(ADP-ribose)-binding promotes Exo1 damage recruitment and suppresses its nuclease activities. DNA Repair (Amsterdam). 35, 106-115 (2015).
- Zhang, F., Shi, J., Chen, S. H., Bian, C., Yu, X. The PIN domain of EXO1 recognizes poly(ADP-ribose) in DNA damage response. Nucleic Acids Research. 43 (22), 10782-10794 (2015).
- Rona, G., et al. PARP1-dependent recruitment of the FBXL10-RNF68-RNF2 ubiquitin ligase to sites of DNA damage controls H2A.Z loading. elife. 7, (2018).
- Young, L. M., et al. TIMELESS forms a complex with PARP1 distinct from its complex with TIPIN and plays a role in the dna damage response. Cell Reports. 13 (3), 451-459 (2015).
- Kong, X., et al. Laser microirradiation to study in vivo cellular responses to simple and complex dna damage. Journal of Visualized Experiments. (131), e56213 (2018).
- Kong, X., et al. Condensin I recruitment to base damage-enriched DNA lesions is modulated by PARP1. PLoS One. 6 (8), 23548 (2011).
- Lan, L., et al. Novel method for site-specific induction of oxidative DNA damage reveals differences in recruitment of repair proteins to heterochromatin and euchromatin. Nucleic Acids Research. 42 (4), 2330-2345 (2014).
- Zerjatke, T., et al. Quantitative cell cycle analysis based on an endogenous all-in-one reporter for cell tracking and classification. Cell Reports. 19 (9), 1953-1966 (2017).
- Ji, Y., Karbaschi, M., Cooke, M. S. Mycoplasma infection of cultured cells induces oxidative stress and attenuates cellular base excision repair activity. Mutation Research. 845, 403054 (2019).
- Sun, G., et al. Mycoplasma pneumoniae infection induces reactive oxygen species and DNA damage in A549 human lung carcinoma cells. Infection and Immunity. 76 (10), 4405-4413 (2008).
- Gassman, N. R., Wilson, S. H. Micro-irradiation tools to visualize base excision repair and single-strand break repair. DNA Repair (Amsterdam). 31, 52-63 (2015).
- Muster, B., Rapp, A., Cardoso, M. C. Systematic analysis of DNA damage induction and DNA repair pathway activation by continuous wave visible light laser micro-irradiation. AIMS Genetics. 4 (1), 47-68 (2017).
- Ikeda, S., et al. Purification and characterization of human NTH1, a homolog of Escherichia coli endonuclease III. Direct identification of Lys-212 as the active nucleophilic residue. Journal of Biological Chemistry. 273 (34), 21585-21593 (1998).
- Rosenquist, T. A., Zharkov, D. O., Grollman, A. P. Cloning and characterization of a mammalian 8-oxoguanine DNA glycosylase. Proceedings of the National Academy of Science U. S. A. 94 (14), 7429-7434 (1997).
- Reid, D. A., et al. Organization and dynamics of the nonhomologous end-joining machinery during DNA double-strand break repair. Proceedings of the National Academy of Science U. S. A. 112 (20), 2575-2584 (2015).
- Taccioli, G. E., et al. Ku80: product of the XRCC5 gene and its role in DNA repair and V(D)J recombination. Science. 265 (5177), 1442-1445 (1994).
- Marsin, S., et al. Role of XRCC1 in the coordination and stimulation of oxidative DNA damage repair initiated by the DNA glycosylase hOGG1. Journal of Biological Chemistry. 278 (45), 44068-44074 (2003).
- Thompson, L. H., Brookman, K. W., Jones, N. J., Allen, S. A., Carrano, A. V. Molecular cloning of the human XRCC1 gene, which corrects defective DNA strand break repair and sister chromatid exchange. Molecular and Cell Biology. 10 (12), 6160-6171 (1990).
- Scharer, O. D.
- Oeck, S., et al. High-throughput evaluation of protein migration and localization after laser micro-irradiation. Science Reports. 9 (1), 3148 (2019).
- Mistrik, M., et al. Cells and stripes: A novel quantitative photo-manipulation technique. Science Reports. 6, 19567 (2016).
- Durand, R. E., Olive, P. L. Cytotoxicity, mutagenicity and dna damage by hoechst 33342. Journal of Histochemistry and Cytochemistry. 30 (2), 111-116 (1982).
- Tobey, R. A., Oishi, N., Crissman, H. A. Cell cycle synchronization: reversible induction of G2 synchrony in cultured rodent and human diploid fibroblasts. Proceedings of the National Academy of Science U. S. A. 87 (13), 5104-5108 (1990).
- Podhorecka, M., Skladanowski, A., Bozko, P. H2AX phosphorylation: Its role in DNA damage response and cancer therapy. Journal of Nucleic Acids. 2010, (2010).
